

Introduction
Amniote tetrapods (i.e., those terrestrial vertebrates that produce eggs in which the embryo is surrounded by a series of extra-embryonic membranes) in the modern world can be divided into two great lines of descent, the Reptilia and the Synapsida. Extant reptiles include lizards, snakes, turtles, the worm-like amphisbaenians, crocodiles, and birds, while monotreme, marsupial, and placental mammals are the extant representatives of Synapsida. The reptile and synapsid lineages both descend from a common ancestor, but that divergence is ancient, occurring in the Carboniferous Period of Earth history (approximately 315 Mya). The first true mammals appear in the fossil record about 200 Mya, near the end of the Triassic Period. If we consider a phylogenetic tree that shows patterns of descent from common ancestors, we can see that there is a large number of extinct members of the synapsid lineage that existed between the origin of Synapsida and the appearance of mammals.
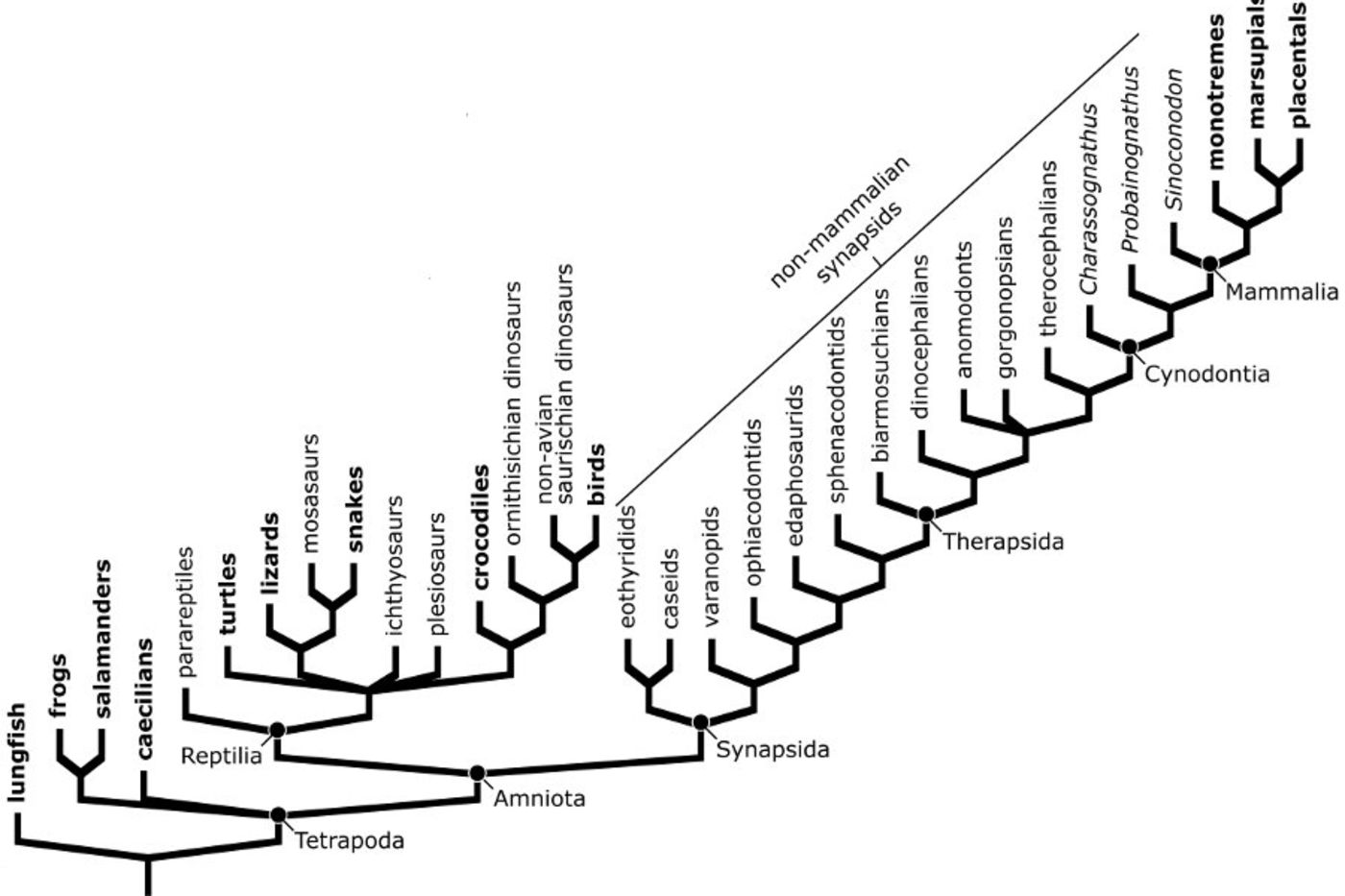
Simplified phylogeny showing relationships among tetrapods. Groups with living members are shown in bold; extinct groups are in plain type. Modified from Angielczyk (2009).
These extinct synapsids are often referred to as “mammal-like reptiles” because some have a superficially reptilian appearance. However, all are descendants of a common ancestor that existed after the divergence between Synapsida and Reptilia, which means they are all more closely related to extant mammals than to any reptile. A more accurate name for these extinct species is “non-mammalian synapsids,” which reflects the fact that they are members of the synapsid lineage, but are not mammals.
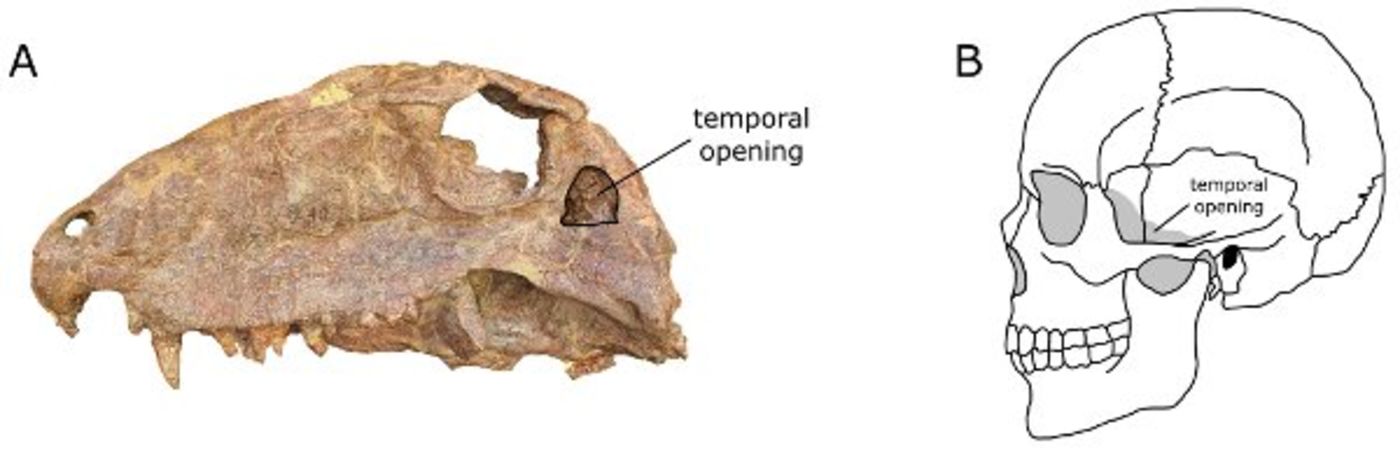
A single temporal opening around which jaw muscles attach is a feature shared by all synapsids. A. location of the temporal opening in the Early Permian synapsid Dimetrodon (FMNH UC 40). B. location of the temporal opening in a human skull. From Angielczyk (2009).
Non-mammalian synapsids are an extremely important part of the fossil record because they document the evolutionary history of many of the distinctive features of mammals, such as the presence of a bony secondary palate, the incorporation of bones from the lower jaw into the middle ear, teeth with complex occlusion patterns, and upright limbs. Morphological features, such as the presence of a single opening behind the eye socket around which jaw musculature attaches, help us recognize members of the synapsid lineage in the fossil record. An introduction to non-mammalian synapsids can be found in Angielczyk (2009).
History of the Collection

Samuel W. Williston in New Mexico, 1911. From Shor (1971).
The history of The Field Museum's non-mammalian synapsid collection begins near the start of the 20th century, with the fieldwork and research of S. W. Williston and P. C. Miller at the University of Chicago, and E. C. Case, a student of Williston who went on to become a professor at the University of Michigan.

Paul C. Miller (left) in New Mexico, 1911. Ermine C. Case (right) in 1891. From Shor (1971).
They made important collections of synapsid fossils from Early Permian-age rocks in Texas, Oklahoma, and New Mexico, and published a seminal series of works describing the specimens and the rocks in which they were found (e.g.,Case, 1907, 1915, 1919; Williston, 1911; Case et al., 1913). The fossils collected by Williston, Case, and Miller became part of the Walker Museum's collections at the University of Chicago. After Williston's death in 1918, A. S. Romer joined the University of Chicago faculty in 1923, and began his extensive fieldwork in the Permian of North America. Between 1923 and 1934, when Romer left for Harvard University, he and P. C. Miller added to the synapsid holdings of the Walker Museum, including an important series of fossils they collected in 1928-1929 in Middle Permian- to Middle Triassic-age rocks in the Karoo Basin of South Africa. Romer was succeeded in 1935 by his student, E. C. Olson. Olson continued the tradition of Permian research at the University of Chicago, and made large collections of synapsids and other vertebrates from Texas and Oklahoma that were deposited in the Walker Museum. Beginning in 1947, the Walker Museum collections were transferred from the University of Chicago to The Field Museum, and Olson continued to deposit specimens at The Field Museum until he left the University of Chicago in 1969.

Left: Alfred S. Romer. Right: Everett C. Olson (far right) instructing University of Chicago students, including future Field Museum Curator John Bolt (far left), in 1963. © The Field Museum GEO82841.
Permian research in Chicago was continued by two University of Chicago students, J. A. Hopson, who joined the University of Chicago faculty in 1967 and whose research focuses on non-mammalian synapsids, and J. R. Bolt, a fossil amphibian specialist who became a curator at The Field Museum in 1972. Former Fossil Mammal Curator J. J. Flynn conducted a series of six expeditions to Madagascar between 1996 and 2003, and added Malagasy Triassic dicynodonts and cynodonts to the non-mammalian synapsid collection. Most recently, K. D. Angielczyk, whose research focuses on anomodont synapsids, joined the curatorial faculty of The Field Museum in 2007. He has collected synapsid specimens from the Permian and Triassic of Tanzania and Zambia, and although the specimens will be returned to their respective countries of origin, casts of important material will remain at The Field Museum.

Left: John J. Flynn with a skull of the cynodont Menadon besairiei from Madagascar. © The Field Museum GEO86240_2c, photographer John Weinstein. Right: Kenneth D. Angielczyk excavates a skull of the anomodont Dicynodontoides nowacki in Tanzania. Photo by R. Smith.
Non-mammalian Synapsid Collection Highlights
The Field Museum's non-mammalian synapsid collection consists of over 1100 catalogued specimens, including 46 holotypes.#
Basal Synapsids
Basal synapsids, often referred to colloquially as “pelycosaurs,” from North America are the major strength of The Field Museum's non-mammalian synapsid collection, comprising approximately 88% of catalogued specimens. The majority of the specimens are from the Lower Permian of Texas and Oklahoma, although some specimens from New Mexico and Illinois also are present.

Composite specimen of Dimetrodon on display in Evolving Planet. Photo by K. Angielczyk.
Members of all major basal synapsid clades are represented, and the collection includes 29 holotypes of caseid, ophiacodontid, edaphosaurid, varanopid, and sphenacodontid species. The collections are historically important, including material collected by WIlliston, Case, Miller, Romer, Olson, and Bolt from many "classic" Lower Permian localities. Nearly complete specimens of Casea broili, Varanops brevirostris, Ophiacodon mirus, and Sphenacodon ferox, as well as composite specimens of Dimetrodon and Edaphosaurs, are on display in the museum's Evolving Planet exhibition.

Skull of the ophiacodontid Varanosaurus sp., from Oklahoma (FMNH PR 1670). Photo by K. Angielczyk.
Olson's North American Therapsids
In a series of works, E. C. Olson described and discussed the evolutionary and biogeographic significance of a number of fragmentary synapsid fossils from the Lower Permian of North America that he suggested represented early members of various lineages within Therapsida (e.g., Olson and Beerbower, 1953; Olson, 1962, 1971, 1974, 1986). Subsequently, these specimens have been reinterpreted as representing more basal synapsids, primarily caseids, varanopids, and sphenacodontids (Sidor and Hopson, 1995; Reisz and Laurin, 2004).

Two of Olson's putative North American therapsids. Left: holotype of Steppesaurus gurleyi, a skull fragment from Texas (FMNH UR 148). Right: a jaw fragment of Eosyodon hudsoni from Texas (FMNH UR 499). Photos by K. Angielczyk.
Nearly all of Olson's putative North American therapsid specimens are housed at The Field Museum, including eight holotypes.
South African Therapsids
The Karoo Basin of South Africa preserves and extremely rich fossil record of Middle Permian- to Middle Triassic-age therapsids that has been studied for over 150 years. In 1928-1929, A. S. Romer and P. C. Miller traveled to the Karoo and made a collection of South African therapsid fossils that is now housed in The Field Museum. The Permian portion of this collection includes five holotypes, and is dominated by dinocephalians (including a nearly complete skull of the rare titanosuchid Jonkeria) and anomodonts (including a skeleton of Aulacephalodon), although gorgonopsians and therocephalians are also present.

Holotype of the Permian gorgonopsian Scymnognathus major from South Africa (FMNH UC 1513). Photo by K. Angielczyk
The Triassic portion of the collection is much smaller, but does include a well-preserved skull of the cynodont Thrinaxodon, a skull of the anomodont Kannemeyeria, and several specimens of the anomdont Lystrosaurus. Specimens of South African dinocephalians, anomodonts, gorgonopsians, and cynodonts can be seen in Evolving Planet.
Malagasy Therapsids
The most recent addition to The Field Museum's non-mammalian synapsid collection is a series of specimens collected by former curator J. J. Flynn and his colleagues in the Triassic of Madagascar. The collection includes several cynodonts, as well as fragmentary material of an anomodont.
References Cited
- Angielczyk, K. D. 2009. Dimetrodon is not a dinosaur: using tree thinking to understand the ancient relatives of mammals and their evolution. Evolution: Education and Outreach 2: 257-271.
- Case, E. C. 1907. Revision of the Pelycosauria of North America. Carnegie Institution of Washington Publication 55: 1-176.
- Case, E. C. 1915. The Permo-Carboniferous red bes of North America and their vertebrate fauna. Carnegie Institution of Washington Publication 207: 1-176.
- Case, E. C. 1919. The environment of vertebrate life in the Late Paleozoic in North America; a paleogeographic study. Carnegie Institution of Washington Publication 283: 1-273.
- Case, E. C., Williston, S. W., and Mehl, M. G. 1913. Permo-Carboniferous vertebrates from New Mexico. Carnegie Institution of Washington Publication 181: 1-81.
- Olson, E. C. 1962. Late Permian terrestrial vertebrates, U.S.A and U.S.S.R. Transactions of the American Philosophical Society (New Series) 52: 1-224.
- Olson, E. C. 1971. Vertebrate Paleozoology. Wiley-Interscience, New York.
- Olson, E. C. 1974. On the Source of therapsids. Annals of the South African Museum 64: 27-46.
- Olson, E. C. 1986. Relationships and ecology of the early terhapsids and their predecessors. Pp. 47-60 in N. Hotton, P. D. MacLean, J. J. Roth, and E. C. Roth, (eds.), The Ecology and Biology of Mammal-like Reptiles. Smithsonian Institution Press, Washington D.C.
- Olson, E. C. and Beerbower, J. R. 1953. The San Angelo Formation, Permian of Texas, and its vertebrates. Journal of Geology 61: 389-423.
- Resiz, R. R. and Laurin, M. 2004. A reevaluation of the enigmatic Permian synapsid Watongia and of its stratigraphic significance. Canadian Journal of Earth Sciences 41: 377-386.
- Shor, E. N. 1971. Fossils and Files. The Life of a Compleat Scientist. Samuel Wendell Williston (1851-1918). Univeristy of Oklahoma Press, Norman.
- Sidor, C. A. and Hopson, J. A. 1995. The taxonomic status of the Upper Permian eotheriodont therapsids of the San Angelo Formattion (Guadalupian), Texas. Journal of Vertebrate Paleontology 15 (supplement to 3): 53A.
- Williston, S. W. 1911. American Permian Vertebrates. University of Chicago Press, Chicago.